NSF:将食品级润滑剂的使用纳入金博宝188官网注册
食品级润滑剂的作用随着食品行业发展成为以高业务量为特点的[详细]
Mettler-Toledo 在中国国际渔业博览会上
Mettler-Toledo 将会在现场展示 X32 高性能 X 射线检测[详细]
ADM 确定了驱动替代蛋白生态系统发展的
正如ADM在2023年全球消费者趋势报告中所强调,随着人们对多[详细]





食品新闻
- 提升全产业链水平,邕之泰致力成为现代一流食品
- 加持县域经济发展 冷链物流如何服务国家战略?
- 长康实业教你如何买到好酱油
- 三只小牛首款NFT「睡眠自由BOX」首发10分钟售罄
- “砥砺奋进 不负韶华” 2022茅台第二届诗歌故
- 步向封控区的夜行者——记疫情下钱大妈的保供
- 道道全-东方山茶油更适合国人的餐桌
- 学会科学减糖:无糖饮料比含糖饮料更好
- 口口椰——健康茶饮开拓者,你喝的每一口都是天
- “喝”走亚健康 轻养良品引领健康饮品新风尚
- 营养抗疫 同心守沪 盼盼食品在行动
- 韦诺(WELRO) NMN成为延缓细胞老化行业市场的标杆
- 2022年西南独特一场展会!聚焦预制食材整场大的
- 超级引擎与中国火箭达成战略合作开启“中国植萃
- 这款冬奥出圈的冻干面,引领方便面食消费新风尚
- 《2022牛排消费趋势洞察》发布,牛肉消费成充满
- 12种口味创新,康师傅对拌面市场再升级!
- 象手食材小剧场打造食材新玩法,助力企业高效拓
- 泉心泉意 晋心晋力 盼盼食品捐赠1100余万抗疫物资
- 赛味馆:Z时代的情绪味觉零食,享受0负担放纵生
- 酒是瓮底香——贵州美酒链赋能新品迭献
- 中国酒业发展研究院正式成立 为中国酒业高质量
- 群星助力集美家居首届抖音短视频大赛暨集美健康
- 果倍爽CapriSun,一推一戳一吸好喝更有FUN
- 酱酒行业乱象降温,郎酒领跑行业健康规范
- 郎酒敢为人先!酱酒行业迎来首个企业内控发布准
- 风靡新茶饮界的舔杯深度钻研细节,"原创手工珍
- 梦龙雪糕生产线迎来新变化,联合利华和路雪太仓
- 热烈祝贺湖北赤壁合作商签约鲜鱼侠!
- 东圣酒业:道酝典范,德厚东圣










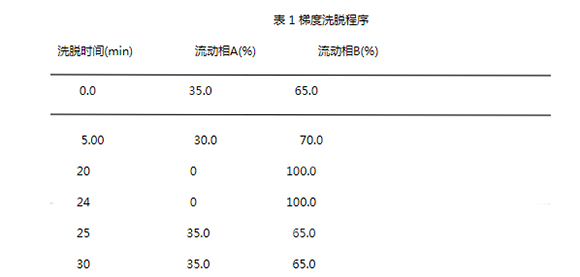












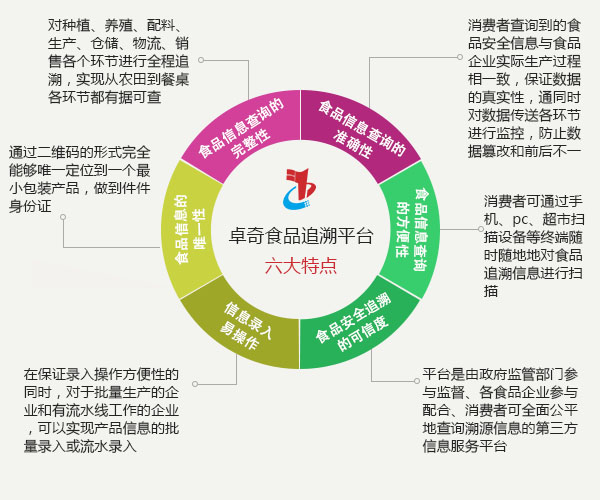

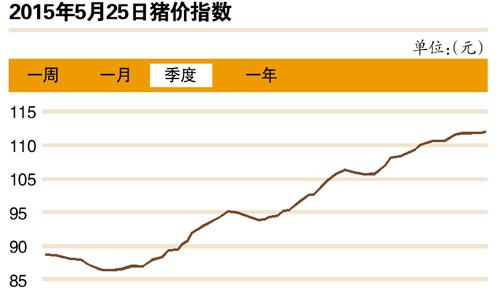















 第十八届中国国际食品加工和包装机械展览会
第十八届中国国际食品加工和包装机械展览会 上海国际食品加工与包装机械展览会
上海国际食品加工与包装机械展览会 第十二届中国食品与农产品安全检测技术与质量控制国际论坛
第十二届中国食品与农产品安全检测技术与质量控制国际论坛 2021亚洲生鲜供应链博览会
2021亚洲生鲜供应链博览会 天然提取物与创新原料展
天然提取物与创新原料展 2023深圳国际营养与健康产业
2023深圳国际营养与健康产业 FBIF2023食品饮料创新论坛
FBIF2023食品饮料创新论坛 第十二届中国国际饮料工业科技展
第十二届中国国际饮料工业科技展 第二十一届中国国际肉类工业展览会
第二十一届中国国际肉类工业展览会 亚食展暨预制食品展
亚食展暨预制食品展 2023第四届内蒙古牛羊产业展览会
2023第四届内蒙古牛羊产业展览会 第二十届北京分析测试学术报告会暨展览会
第二十届北京分析测试学术报告会暨展览会 2023中国营养及功能食品峰会
2023中国营养及功能食品峰会 第二十一届中国国际科学仪器及实验室装备展览会
第二十一届中国国际科学仪器及实验室装备展览会 慕尼黑上海分析生化展
慕尼黑上海分析生化展
 深圳国际预制菜暨食品供应链博览会
深圳国际预制菜暨食品供应链博览会 2023生物发酵展(上海)
2023生物发酵展(上海) 上海国际食品加工与包装机械联展
上海国际食品加工与包装机械联展 第二十四届健康天然原料、食品配料中国展
第二十四届健康天然原料、食品配料中国展 第二十四届健康天然原料、食品配料中国展
第二十四届健康天然原料、食品配料中国展 第十三届中国国际健康产品展览会
第十三届中国国际健康产品展览会 中国成都国际分析测试与实验室技术设备博览会
中国成都国际分析测试与实验室技术设备博览会 2024第13届中国西安糖酒食品交易会
2024第13届中国西安糖酒食品交易会 2023中国制冷展
2023中国制冷展 第十四届深圳国际营养与健康产业展
第十四届深圳国际营养与健康产业展 第六届中国教育后勤展览会
第六届中国教育后勤展览会